AAPs Rule!
Current projects:
Description
Carbon is the most actively cycled element in the biosphere. Comprehending the carbon cycle is crucial for understanding biological processes. The natural carbon cycle has been imbalanced by anthropogenic activities, resulting in the climate change, and affecting not only wild life, but also human health and well-being [1, 2]. Freshwater lakes are a significant component of the global carbon cycle: they cover only about 3% of Earth’s surface [3], but they bury annually more organic carbon in their sediments than the oceans [4]. Still, they are a net source of CO2 to the atmosphere [5]. Lakes are under substantial anthropogenic pressure, and their contribution to the carbon cycle is changing [4]. Therefore, understating the carbon cycle in lakes is crucial for understanding the global carbon cycle, and forecasting the climate change.![]()
Microbes are the key component of the carbon cycle in aquatic habitats [6]. Eukaryotic algae and cyanobacteria fix CO2 during photosynthesis, leading to the produced organic matter being recycled back to CO2, mainly during microbial respiration [7, 8]. Rates of microbial heterotrophic activities, namely bacterial respiration and production, are considered to be independent of light, and consequently, they are measured in the dark [9-11]. These values are then used to calculate the efficiency of conversion of organic matter into microbial biomass (bacterial growth efficiency): 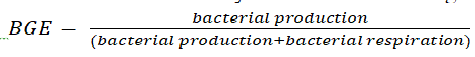 .
.
BGE controls flows of energy and matter to higher trophic levels, and the balance between the light-dependent primary production and the light-independent respiration (that is, net phototrophy or net heterotrophy) in the ecosystem [12, 13].
Discovery that photoheterotrophic bacteria are abundant in the ocean questioned the assumption that in situ microbial respiration and production occur at the same rates under light and in the dark [14, 15]. Two groups of photoheterotrophic bacteria thrive in aquatic environments: rhodopsin containing bacteria [14], and aerobic anoxygenic phototrophic (AAP) bacteria [15]. Rhodopsin is a retinal-binding membrane protein that acts as a light driven proton pump. Rhodopsin containing bacteria include the most abundant but less active and slowly growing freshwater taxa, like Actinobacteria and LD12 Alphaproteobacteria [16-19]. AAP bacteria use bacteriochlorophyll-containing reaction centres to harvest energy from light. Freshwater AAP bacteria belong mainly to Betaproteobacteria (Limnohabitans, Polynucleobacter) and Alphaproteobacteria (Sphingomonas, Methylobacterium) [20-22]. Abundance of AAP bacteria varies seasonally, and they contribute up to 30% of total bacterial abundance during maxima in spring and late summer [23-25]. They exhibit larger cell size than an average bacterium, higher activity than an average bacterium, fast growth rates, and high susceptibility to grazing [19, 23, 26, 27]. Experiments with pure cultures showed that AAP bacteria decrease their respiration by 75%, and increase growth efficiency by 50% when grown in light:dark cycles compared to the dark [28]. It has been hypothesised that thanks to this additional energy, AAP bacteria have an advantage over heterotrophic bacteria under carbon limitation, and that they can access refractory organic carbon (rDOC), that is unavailable to heterotrophic bacteria [29]. However, the in situ benefits of light harvesting by AAP bacteria and their dynamics, activity and growth rates are still not well understood [30]. Taken together, the role of AAP bacteria in the carbon cycle remains an open question [29-31].
Despite the fact that information on AAP bacteria accumulates, and that the potential importance of photoheterotrophy has been realized almost two decades ago, rates of bacterial respiration and production are still typically measured in the dark [30]. This can substantially bias the estimates of bacterial growth efficiency in times of elevated abundance of photoheterotrophs, especially of highly active AAP bacteria [19, 32].
Here, we propose a novel approach to address the question of contribution of photoheterotrophy to the freshwater carbon cycle. We will compare in situ rates of microbial respiration and production measured in the dark and under infrared light (λ=850 nm). Infrared (IR) is absorbed only by bacteriochlorophyll-containing AAP bacteria, therefore our measurements will not be affected by photosynthetic algal activity. Moreover, to understand the expected temporal variability in contribution of photoheterotrophy to the carbon cycle in lakes [30, 33], and to enhance knowledge on ecology of AAP bacteria, we will study their diversity, growth rates, and in situ dynamics and activity.
References
[1]: Writing Core Team, IPCC, 2014: Climate Change 2014: Synthesis Report. R Pachauri and L Meyer [eds] pp. 151. [2]: Pecl et al., 2017, Science 355: eaai9214. [3]: Downing et al., 2006, Limnol Oceanogr 51:2388. [4]: Tranvik et al., 2009, Limnol Oceanogr 54:2298. [5]: Sobek et al., 2005, Global Biogeochem Cycles 19: GB2003. [6]: Canfield et al., 2005 Heterotrophic Carbon Metabolism, in Adv Mar Biol, DE Canfield, E Kristensen, and B Thamdrup [eds] pp. 129-166. [7]: Del Giorgio et al., 1999, Ecology 80:1422. [8]: Jansson et al., 2000, Ecology 81:3250. [9]: Kirchman et al., 1985, Appl Environ Microbiol 49:599. [10]: Fuhrman & Azam, 1982, Mar Biol 66:109, [11]: del Giorgio & Duarte 2002, Nature 420:379. [12]: Sinsabaugh et al., 2013, Ecol Lett 16:930. [13]: del Giorgio 1998, Annu Rev Ecol Syst 29: p503. [14]: Beja et al., 2000, Science 289:1902. [15]: Kolber et al., 2001, Science 292:2492. [16]: Salcher et al., 2013, Isme J 7: 896. [17]: Šimek et al., 2006, Environ Microbiol 8:1613. [18]: Glockner et al., 2000, Appl Environ Microbiol 66:5053. [19]: Ferrera et al., 2011, Appl Environ Microbiol 77:7451. [20]: Salka et al., 2011, Environ Microbiol 13: p. 2865. [21]: Zeng et al., 2012, J Bacteriol 194:6302 [22]: Martinez-Garcia et al., 2012, ISME J 6:113. [23]: Cepáková et al., 2016, Environ Microbiol 18:5063. [24]: Cuperova et al., 2013, Appl Environ Microbiol 79:6439. [25]: Lew et al., 2015, Folia Microbiol 60:165’ [26]: Masin et al, 2008, Environ Microbiol 10:1988. [27]: Garcia-Chaves et al., 2016, Isme J 10:1579, [28]: Hauruseu & Koblížek, 2012, Appl Environ Microbiol 78:7414. [29]: Koblížek 2015, FEMS Microbiol Rev, 39:854. [30]: Ruiz-González et al., 2013, Front Microbiol 4:131. [31]: Gasol et al., 2008, Aquat Microb Ecol 53:21. [32]: Salcher 2014, J Limnol 73:74. [33]: Baptista et al., 2014, Aquat Ecol 48:327
Team:
Dr. Katarzyna Piwosz (PI)
MSc. Izabela Mujakić
MSc. Vesna Grujčić
Ing. Martina Hanusová
Jason Dean BSc.
International collaborators:
Dr. Michaela M. Salcher, Limnological Station, the University of Zurich, Switzerland
Dr. Jürgen Tomasch, Helmholz Centre for Infection Research in Braunschweig, Germany